
|
Our Valuable Native Grasslands, Better Pastures Naturally Proceedings of the Second National Conference of the Native Grasses Association |


Vegetation changes in southeast australian temperate grasslands
Denys Garden1 and Terry Bolger2
1NSW Agriculture, 2CSIRO Plant Industry
GPO Box 1600, Canberra ACT 2601
Introduction
Because of isolation, unique vegetation originally developed in Australian grasslands. This vegetation was very sensitive to changes in grazing, soil fertility and the introduction of exotic species. The original vegetation was characterised by a dominance of perennial grasses. There were few legumes present and, consequently, soil N levels were likely to have been low. After settlement, livestock and agricultural practices were imported from Europe. The effects of grazing by domestic and feral animals, accidental introduction of cool-season annual grasses, legumes and forbs, and a changed pattern of burning were marked. In addition, exotic plants were deliberately introduced into agricultural areas, often with cultivation. This paper seeks to describe the botanical changes that have occurred and to explain them in terms of recently proposed ecological processes.
Patterns of change
Botanical change in the temperate grasslands of south-east Australia has been described by Moore (1993). Figure 1 shows the pathways of change for pastures on the southern tablelands of NSW that were either originally unsown, or cultivated and sown. The first major change in composition (Stage I to Stage II - Figure 1) was from taller warm-season perennial grasses (e.g. Themeda australis) to shorter cool-season perennials such as Austrodanthonia spp. and Austrostipa spp. Further grazing and the application of fertiliser accelerated these changes, moving composition from Stage II to Stage III. Legumes and annual grasses increased as a result of increased soil fertility. While Austrostipa spp. remained, the balance of native perennial grasses shifted more towards Austrodanthonia spp. Further degeneration of pastures to the final stage of annual dominance described by Moore (1993) is indicated by the path from III to IV. Although Figure 1 allows for the possibility of reversion from Stage IV to III, the mechanisms are unclear, and it is unlikely to be easily achieved. Stage IV is an inherently unstable system, as changes in botanical composition are unpredictable and reliant on timing of rainfall events. Therefore, it is important that the perennial grass content of grasslands is preserved
Where soils are cultivated, and introduced species are sown with fertiliser, grasslands from Stages I-IV (Figure 1) can be converted to Stage V. Depending on soil conditions (e.g. soil acidity) and the success of establishment, resultant pastures can range from highly successful sown pastures to those with only low proportions of exotic species present. Also, composition can move from Stage V to VI under the influence of increased soil acidity, reduced fertiliser, drought and overgrazing, a process often referred to as `pasture decline'. Stage VI pastures are common on the tablelands of NSW. While there are some remaining introduced perennial grasses, the proportion is low, and native year-long green grasses (e.g. Austrodanthonia spp.) are able to re-establish. Depending on season and fertiliser, inter-tussock spaces are filled with introduced cool-season annual grasses, legumes and weeds. Such pastures are stable and productive under a wide range of conditions, providing the perennial:annual ratio is high (Garden et al. 2000).
Further decline (from Stage VI back to IV) results when a pasture is destabilised, perennial grasses are reduced, and bare areas are created which allow recruitment sites for annual grasses. This situation can be created by overgrazing during drought, and/or by high stocking rates. As described earlier, the annual dominant Stage IV state is an inherently unstable one.
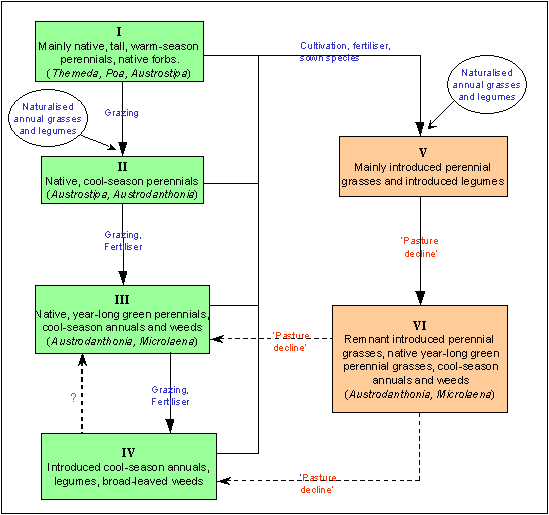
Figure 1. Sequence of botanical changes in native grasslands on the southern tablelands of New South Wales, with (right hand side) and without (left hand side) cultivation and sowing of exotic species. Species listed are dominants only.
Ecological processes underlying botanical change
Recently, the roles of two complementary ecological processes, competition and recruitment limitation (the inability of species to disperse, germinate or establish successfully), have been recognised as important determinants of plant community structure (e.g. Tilman 1997). We propose that these processes may have been important in determining change in Australian temperate grasslands, through interactions with grazing and other management and environmental factors. Studies in the North American prairies show that nitrogen (N) is often the most limiting resource in many grasslands (Wedin 1995), and that the competitive dominants are likely to be the species that can reduce the concentration of the limiting resource to the lowest level (Wedin and Tilman 1993). The original dominants of Australian temperate grasslands warm-season perennial grasses generally good competitors for N and tend to maintain soil mineral N at low levels (Moore 1993). Thus, these species can create the low N conditions in which they are superior competitors. This positive feedback between plant competition and N cycling may be an important process in maintaining the stability of many grassland plant communities, including the Stage I grasslands referred to in Figure 1.
Grazing precipitated the change from Stage I to Stage II pastures (Figure 1). The native, warm-season perennials were poorly adapted to grazing and, thus, grazing reduced the cover and production of these dominants, disturbed the soil surface creating bare sites for invasion of other species, and increased the rate of nutrient cycling via the animal pathway. Under this new regime, the previously subordinate cool-season perennial grasses, which were more tolerant of grazing and better recruiters, were able to dominate. Also, Stage II pastures had higher levels of soil mineral N, allowing naturalised annual species to invade.
Greater fluctuations in soil mineral N under Stage III pastures, in combination with climatic variation, can result in large fluctuations in the proportions of perennials and annuals within and across years. These transient and reversible fluctuations in species composition generally do not cross thresholds and, thus, these Stage III pastures are considered to be stable under a wide range of conditions (Garden et al. 2000). However, continued grazing and inputs of fertiliser, perhaps in combination with climatic events such as drought, can destabilise these Stage III pastures and shift them to a state dominated by less stable and less productive annual species (Stage IV).
These ecological processes are hypothesised to operate in a similar manner in determining species composition in cultivated situations, as outlined in Figure 1. The main difference is that the introduced perennial grasses are poor recruiters, so changes in pasture states (Stage V to Stage VI, and Stage VI to Stage III or IV) due to loss of these species are considered essentially irreversible, due to recruitment limitation. This could explain the large number of previously sown pastures on the tablelands of NSW now dominated by a combination of native perennial and exotic annual grasses. The importance of the complementary roles of competition and recruitment limitation in structuring such diverse plant communities as tallgrass prairie and tropical forests suggests that these processes are generic and likely to be important in Australian temperate grasslands as well.
References
- Garden, D.L., Lodge G.M., Friend D.A., Dowling P.M. and Orchard, B.A. (2000). Effects of grazing management on botanical composition of native grass-based pastures in temperate southeast Australia. Australian Journal of Experimental Agriculture 40: 225-245.
- Moore, R.M. (1993). Grasslands of Australia. Ch. 13. In, "Ecosystems of the world. 8B. Natural Grasslands. Eastern hemisphere and résumé". pp. 315-360. Ed. R.T. Coupland. (Elsevier, Amsterdam).
- Tilman, D. (1997). Community invasibility, recruitment limitation, and grassland biodiversity. Ecology 78: 81-92.
- Wedin, D.A. (1995). Species, nitrogen and grassland dynamics: the constraints of stuff. Ch. 24. In, `Linking species and ecosystems'. pp. 253-262. Eds. C.G. Jones and J.H. Lawton. (Chapman and Hall, New York).
- Wedin, D.A. and Tilman, D. (1993). Competition among grasses along a nitrogen gradient: initial conditions and mechanisms of competition. Ecological Monographs 63: 199-229.
